Примечание: пунктом последней репродукции сортообразцов была Адлерская опытная станция ВИР – (АОС).
Интродукционные испытания пяти сортообразцов V. unquiculata в 2008 – 2009 гг. проводились на двух (50 и 1830 м) высотных уровнях Дагестана. Посев семян был проведён в метровых рядах с расстоянием между ними 20 см. Однако в условиях Внутреннегорного Дагестана (окрестности селения Шитли Гунибского района, 1830 м над ур. м., юго-западная экспозиция склона) посевы этой тёплолюбивой культуры положительных результатов не дали. Семена всех пяти сортообразцов V. unquiculata не взошли. Отрицательные результаты были получены в пределах такого же высотного уровня и ранее (1999 г.) при интродукции в условиях Гунибской экспериментальной базы (1950 м над ур. м., северный склон) Горного ботанического сада ДНЦ РАН. Видимо, определяющую роль в этом сыграли природные факторы, обусловленные высотой 1000 м над ур. м. и более. В районах Внутреннегорного Дагестана для выращивания вигны китайской не хватало необходимого тепла. В условиях же Низменного Дагестана (Кумторкалинский р-н, зимнее пастбище Гунибского р-на, урочище Хумтуп, 50 м над ур. м., с. ш.– 43º02′45′′ и в.д. – 47º13′50′′) в первом же году испытания все сортообразцы дали дружные ювенильные растения в достаточном количестве – более 20 особей (рис. 2). В процессе роста и развития проводились фенологические наблюдения. 
Рис. 2 – Ювенильные растения сортообразцов V. unquiculata, полученные в условиях равнинного Дагестана После завершения вегетационного цикла у 20 генеративных побегов каждого сортообразца, представляющих надземную часть растения, были учтены 12 признаков, которые нами были отнесены к двум группам: морфологическим (размерным и числовым) и весовым. В первый год (2008 г.) испытаний в условиях Низменного Дагестана у интродуцентов было получено недостаточное количество растений сортообразцов, поэтому то в настоящем сообщении в общих чертах описывается только изменчивость четырёх весовых признаков растений-культиваров, полученных на второй год выращивания (2009 г.). Кроме того, дополнительно были вычислены признаки адаптивной стратегии – репродуктивное усилие (Re) и для средних величин – эффективность его (Eff ). Последние признаки относятся к характеристикам семенной продуктивности и, являясь главными показателями адаптивной стратегии, показывают конкретную долю, выделяемую на размножение. Работа выполнена на популяционном уровне. Статистическая обработка данных проводилась в соответствии с общепринятыми методами, были получены результаты как суммарной статистики, так и корреляционного и дисперсионного анализов [8, 9]. Также дополнительно для средних величин признака были вычислены значения t-критерия Стьюдента. При проведении части расчётов использовали ПСП Statgraf, version 3.0. Sharevarе, система анализа данных Statistica 5.5. Некоторые предварительные результаты вариабельности учтённых признаков объединённой выборки (n = 100) культиваров V. unquiculata в условиях Равнинного Дагестана нами были представлены ранее [10]. Результаты исследования. При сравнительном анализе структуры изменчивости средних значений весовых признаков генеративного побега, обычно представляющего надземную часть у культурных травянистых однолетних растений, объединённой выборки (n = 100) и сортообразцов V. unguiculata выяснилось, что для всех рассматриваемых здесь четырёх весовых признаков (X – сухая масса стебля, X1 – сухая масса плодов, X2 – сухая масса семян и X3 – сухая масса генеративного побега) и совокупности особей характерны высокие, максимальные, сравнительно сходные значения главного показателя вариабельности – коэффициента вариации (табл. 2). 2. Сравнительная характеристика изменчивости весовых признаков сортов V. unguiculata в условиях Низменного Дагестана (50 м высоты над ур. м.), n = 100
Примечание (здесь и далее): X – сухая масса стебля, X1 – плодов, X2 – семян, X3 – генеративного побега, Re – репродуктивное усилие.
Среди этой группы признаков при максимальном значении последнего показателя (120,0 %) выделяется наиболее пластичный признак – сухая масса стебля (X), для которого отмечен минимальный показатель амплитуды крайних величин (Max – Min = 173,58). Подобные высокие показатели относительной изменчивости этих признаков объединённой выборки (n = 100) указывают на совокупность данных из разных групп интродуцентов. В то же время максимальный показатель размаха крайних вариантов (Max – Min) свойственен сухой массе генеративного побега (X3) и разница равна (476,83 – 2,2) = 474,63. При этом для выборок всех четырёх учтённых здесь признаков сухой массы отмечены, как и следовало бы ожидать, сравнительно высокие показатели относительной изменчивости (56,9 – 110,8 %). Рассматриваемые сортообразцы различались также по колебаниям средних значений весовых признаков. Если для средних показателей сухой массы стебля (X) характерен минимальный (47,5 – 10,9) = 36,6 мг размах, то для средних показателей сухой массы генеративного побега (X3) отмечена максимальная амплитуда (125,9 – 37,4) = 88,5 мг. Два других весовых признака, относящихся к показателям семенной продуктивности, в этом отношении занимают промежуточное положение, и величины разницы средних значений сухой массы плодов (X1) и семян (X2) крайних сортообразцов сравнительно близки друг к другу и равны (78,4 – 22,2) = 56,2 и (63,5 – 16,7) = 46,8 мг соответственно. Однако из этой группы своим сравнительно низким показателем относительной изменчивости (45,5 %) выделяется относительный признак генеративной сферы – репродуктивное усилие (Re), который является главным показателем адаптивной (репродуктивной) стратегии и показывает долю (62,7 %), выделяемую организмом на репродукцию. Для этого относительного признака, в отличие от учтённых весовых признаков, отмечена, хотя и несущественная, максимальная величина отрицательной корреляционной связи (rxy = – 0,791) между средними значениями признаков и их коэффициентами вариации. Недостоверность корреляции, на наш взгляд, связана с числом степеней свободы (df = n – 2 = 3), которое зависит от числа выборок. Для того чтобы данная величина связи была значимой необходимо изучить как минимум 8 сортообразцов данной культуры [8]. Среди интродуцированных в условиях Низменного Дагестана пяти сортообразцов V. unquiculata максимальными средними показателями весовых признаков, особенно показателями семенной продуктивности и репродуктивного усилия (Re), выделяется культивар № 133 ВИР селекции Ростовской области. При этом минимальные величины этих признаков присущи интродуценту № 134, выведенному украинскими селекционерами. В то же время минимальные средние показатели признаков семенной продуктивности (X1 и X2) и доли, выделяемой растением на репродукцию (Re = 0,506209), отмечены для сортообразца № 141 селекции Китайской Народной Республики. Средние величины учтённых здесь весовых признаков других культиваров занимают промежуточное положение. Однако крайние варианты минимальных и максимальных средних показателей эффективности репродуктивного усилия – Eff и доли, выделяемой растением на репродукцию (Re), не совпадают (рис. 3). Эти относительные признаки – Re и Eff относятся к показателям адаптивной (репродуктивной) стратегии. 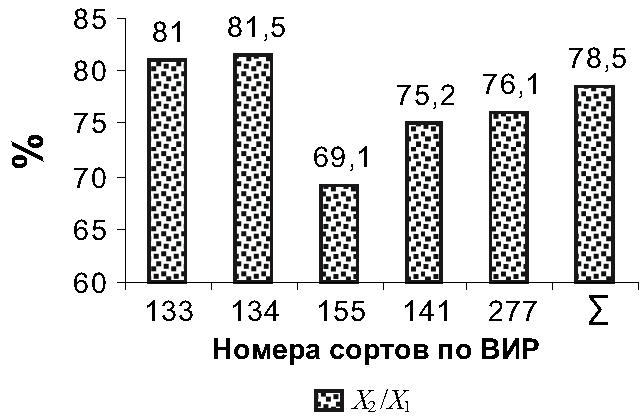
Рис. 3 – Эффективность репродуктивного усилия (Eff ) – доля (%) сухой массы семян таковой в бобах (X2/X1) сортообразцов V. unguiculata в условиях Низменного Дагестана Если репродуктивное усилие (Re) показывает долю, выделяемую на репродукцию генеративным побегом или растением в целом, то эффективность репродуктивного усилия (Eff ) – отношение сухой массы семян к таковой бобов [11] отражает конкретный процент, ассигнуемый генеративной сферой (соцветием, плодом) на размножение. Наибольшая доля (81,5 %) сухой массы семян таковой в бобах (X2/X1) присуща культивару № 134, у которого отмечены минимальные показатели сухой массы стебля (X) и генеративного побега (X3). Минимальная же порция (69,1 %) сухой массы семян (X2/X1) в плодах характерна интродуценту № 155 американской селекции, а остальная доля в сухой массе плода (X1) приходится на сухую массу створок боба. При сравнении средних показателей весовых признаков сортообразцов V. unquiculata по t-критерию Стьюдента выяснилось, что средние величины всех четырёх учтённых весовых величин сортообразцов № 133 и 134 различаются существенно – на 99,0- и 99,9%-ном уровнях достоверности (табл. 3). 3. Сравнительная характеристика средних значений весовых признаков растений сортообразцов V. unquiculata по t-критерию Стьюдента (n = 20) (df = n1 + n2 – 2 = 38)
Примечание: t – критерий Стьюдента, df – число степеней свободы. Прочерк означает отсутствие существенного различия. * Р < 0,05; ** Р < 0,01; *** P < 0,001. Кроме того, средние показатели сортообразца № 133 V. unquiculata селекции Ростовской области в большинстве случаев заметно отличаются по данному критерию от остальных культиваров. Различия же средних показателей всех четырёх весовых признаков (X, X1, X2 и X3) четырёх (№ 134, 141, 155 и 227) интродуцентов между собой не существенны и носят случайный характер. При этом сравнительно близки и сходны средние величины относительного признака – репродуктивного усилия (Re) интродуцентов, который в большей мере контролируется генотипом, и различия между всеми вариантами сравнения средних значений по t-критерию Стьюдента также не значимы. Кроме того, в преобладающем большинстве случаев между весовыми признаками наблюдаются существенные значения положительной корреляционной связи (табл. 4). При этом между весовыми признаками двух вариантов – между сухой массой плодов (X1), с одной стороны, и сухой массой семян (X2) и генеративного побега (X3), с другой, всех сортообразцов и объединённой выборки (n = 100) также отмечены существенные, на самом высоком (99,9 %) уровне достоверности, положительные корреляционные связи. 4. Сравнительная характеристика корреляционных связей (rxy) весовых признаков сортообразцов V. unquiculata при интродукции в условиях Низменного Дагестана (n = 20), (df = n – 2)
Примечание: n – объём выборки; ∑ – объединённая выборка; df – число степеней свободы. Значения коэффициентов корреляции (rxy) приведены в виде первых двух знаков после запятой. Прочерк означает отсутствие существенной связи. * Р < 0,05; ** Р < 0,01; *** P < 0,001.
Между сухой массой семян (X2) и сухой массой генеративного побега (X3) также замечены крепкие значимые корреляции. Иначе говоря, между тремя весовыми признаками – сухой массы плодов (X1), семян (X2) и генеративного побега (X3), как и следовало бы ожидать, во всех вариантах существуют прочные связи. Кроме того, сухая масса стебля (X) с высокой степенью имеет и крепкие корреляции с остальными тремя весовыми (X1, X2 и X3) признаками всех интродуцентов, за исключением одного варианта. Эти весовые признаки в преобладающем большинстве случаев положительно коррелированы с долей (Re), выделяемой генеративным побегом на размножение. В результате проведённого однофакторного дисперсионного анализа выяснилось, что учтённый фактор – разнообразие сортового материала относительно одинаково и существенно на 99,0%-ном уровне достоверности влияет на изменчивость всех учтённых весовых признаков сортообразцов новой для Дагестана культуры – V. unquiculata и сила влияния (h2, %) колеблется в пределах от 22,4 до 24,2 % (табл. 5). Однако на вариабельность относительного признака – репродуктивного усилия (Re), который намного крепче контролируется генотипом, сортовое разнообразие заметного влияния не оказывает, и оно носит случайный характер. 5. Результаты однофакторного (сортовое разнообразие) дисперсионного анализа изменчивости весовых признаков сортообразцов V. unquiculata (n = 100)
Примечание: mS – дисперсия. F – критерий Фишера. В скобках указано число степеней свободы (df). h2 – сила влияния фактора. Тире означает отсутствие существенного влияния фактора. * Р < 0,05; ** Р < 0,01; *** Р < 0,001.
Выводы. Впервые в Низменном Дагестане (50 м над ур. м.) успешно прошло интродукционное испытание сортообразцов новой и нетрадиционной для Дагестана культуры из Африки – коровьего гороха – V. unquiculata. Из интродуцированных культиваров выделен с максимальными средними показателями учтённых весовых признаков сортообразец № 133 (по каталогу ВИР) селекции Ростовской области, а с минимальными (в преобладающем большинстве случаев) – № 134 украинской селекции. Установлено, что на изменчивость учтённых весовых признаков существенно и в одинаковой степени влияет фактор сортовое разнообразие, при этом сила влияния (h2, %) колеблется от 22,4 до 24,2 %. Сортовое разнообразие недостоверно влияет на изменчивость главного показателя адаптивной (репродуктивной) стратегии – репродуктивное усилие, а различия средних показателей по t-критерию Стьюдента носят случайный характер. Высокая доля семян в плоде свидетельствует об эффективности репродуктивного усилия, и в пределах культиваров она колеблется от 69,1 до 81,5 %. Литература 1. Мамаев С.А. О проблемах и методах внутривидовой систематики древесных растений. Амплитуда изменчивости // Закономерности формообразования и дифференциации вида у древесных растений. Свердловск, 1975. С. 3 – 38. 2. Марков М.В., Плещинская Е.Н. Влияние фенотипической среды на проявление признаков жизненной формы и репродуктивной стратегии у растений // Взаимодействие между компонентами экологических систем. Казань: Изд-во Казан. ун-та, 1985. С. 99 –108. 3. Магомедмирзаев М.М., Гусейнова З.А. Об адаптивных стратегиях интродуцируемых видов окультуренной люцерны // Интродукционные ресурсы горного растениеводства. Махачкала, 1996. С. 120 – 132. 4. Драгавцев В.А., Острикова В.М. Клинальные модели растительных популяций и метод оценки уровней механизма акклиматизации // Генетика. 1966. Т. 2. № 3. С. 34 – 44. 5. Синская Е.Н. Проблема популяций у высших растений. Л.: Сельхозиздат, 1961. С. 3 – 69. 6. Сельскохозяйственный энциклопедический словарь / гл. ред. В.К. Месяц. М.: Советская энциклопедия, 1989. 656 с. 7. Биология. Большой энциклопедический словарь / гл. ред. М.С. Гиляров. М.: Советская энциклопедия, 2001. 94 с. 8. Зайцев Г.Н. Методика биометрических расчётов. М.: Наука, 1983. 256 с. 9. Лакин Г.Ф. Биометрия. М.: Высшая школа, 1990. 352 с. 10. Шуайбова Н.Ш., Хабибов А.Д., Омарова П.А. К оценке показателей вариабельности культиваров Vigna unguiculata (L.) WARP. в Равнинном Дагестане // Биологическое разнообразие Кавказа и Юга России: матер. XXI Междунар. науч. конф. (г. Магас, 15 – 18 ноября 2019 г.). Магас: ООО «КЕП», 2019. С. 256 – 259. 11. Магомедмирзаев М.М. Введение в количественную морфогенетику. М.: Наука,1990. 230 с. DOI 10.37670/2073-0853-2020-82-2-81-87
|
Для авторов2025 - Год защитника Отечества
Экология Оренбуржья
Оренбуржье: события и факты
Страничка библиографаОформление списка
Внешний электронный ресурс
СтатистикаПросмотры материалов : 1885936 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 № 2 (82)
№ 2 (82)








